- Complexes of lipids and DNA molecules
Somatic gene therapy holds great promise for future medical applications including, for example, novel treatments for various inherited diseases and cancers. Complexes composed of cationic lipids (CLs) and DNA, named lipoplexes, constitute one of the most promising nonviral gene delivery systems. They have attracted considerable attention due to their inherent advantages over viral delivery methods. These advantages include simple and variable preparation, unlimited length of the transported DNA, and lack of a specific immune response due to the absence of viral peptides and proteins. However, their gene transfer efficiency is currently considerably lower than that of viral vectors and substantial improvements are required before CL-DNA complexes will be viable for therapeutic purposes.
The improvement of lipid vectors requires a better understanding of their mechanism of transfection, and the chemical and physical parameters of CL-DNA complexes that influence it. To gain insight into the biophysical behavior of these complexes, we developed computer models that allow the study of molecular self-assembly from structural disorder. Computational simplifications necessary for efficiency are introduced through a coarse-grained representation of the intra-molecular atomic details. The inter-molecular potentials are designed to mimic the hydrophobic effect without the explicit presence of solvent. Thus, the approach carefully balances the need for molecular detail with computational practicality in a manner that allows for solvent-free simulations of complex self-assembly over long enough time scales to address experimental reality. Figure A shows one of the structures observed in our simulations - a novel phase (which had not been observed experimentally yet), where DNA rods and cylindrical micelles form a 2D lattice analogous to the 3D Na-Cl-type structure.
In addition to showing spontaneous self-assembly of CL-DNA complexes, the broad utility of the model has been illustrated by demonstrating excellent agreement with X-ray diffraction experimental data. We studied the structural and thermodynamic properties of lamellar complexes containing both monovalent and multivalent CLs. In the latter case, the condensation of the DNA molecules is greatly enhanced by attractive, CL-mediated, DNA-DNA interactions. Figure B illustrates the different compaction regimes of complexes of multivalent lipids, as a function of the area charge density of the membranes (which can be varied by adding different amounts of neutral lipids to the system). Examining published transfection efficiency (TE) data in the light of our results supports a previously proposed hypothesis that stability and TE of CL-DNA complexes are oppositely correlated.
(A)

(B)
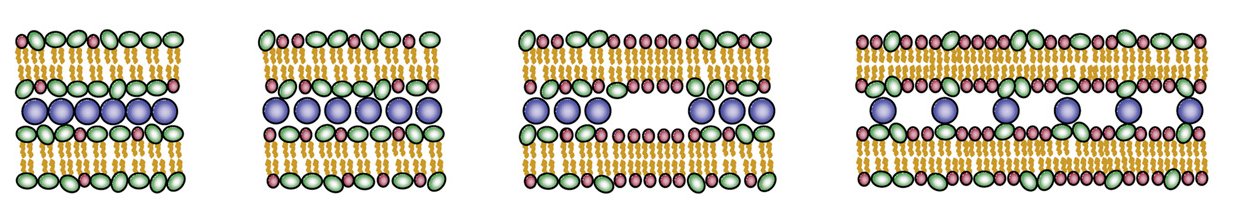
Figure: (A) A fully charged complex (consisting of CLs only) with a square lattice arrangement of DNA rods (shown in blue) and cylindrical micelles. Each charged lipid is represented as a trimer consisting of two hydrophobic beads (white) and one hydrophilic charged bead (red). (B) Different compaction regimes of lamellar complexes of multivalent lipids (from left to right): i - high charge density close packed structured, ii - lower charge density condensed, but not close-packed, state, iii - further reducing the charge density leads to phase separation between the condensed complex and regions of neutral membrane, iv - low charge density expanded state.
- Adhesion domains in supported and decorated membranes
Supported membranes with reconstituted proteins ("decorated") are designed to mimic natural biomembranes. They offer a framework for probing certain cellular processes and may serve as the basis for new biomimetic devices. In recent years, there has been an enormous progress in the development of advanced fabrication and characterization techniques of supported and decorated membranes. Our research effort aims to improve the theoretical understanding of such systems by investigating their structural and mechanical properties, as well as their dynamical behavior. This is done using a combination of analytical models, solvent-free coarse-grained molecular simulations, as well as statistical mechanical analysis of lattice gas models. We are especially interested in the formation of adhesion domains, i.e., regions enriched in bonds that connect the membrane to the surface. The figure below shows two equilibrium configurations taken from our lattice-gas simulations. In both figures we consider a dilute lattice-gas (fraction of occupied lattice sites is 0.1) with relatively weak attractive attraction between the sites (the nearest-neighbor interaction energy is kT). Figure A corresponds to the standard-lattice gas model which, under these conditions, is in the gas phase. In Figure B we include an addition energy term representing the membrane-mediated interactions between the adhesion bonds which are induced by the thermal undulations of the membrane. With this additional interaction energy term, the system undergoes a phase transition into the condensed phase and a compact adhesion cluster is formed in the membrane.
(A)
(B)

Figure: Equilibrium configuration of a membrane consisting of 2000 lipids that fluctuates above a plane surface (frame indicated by a thick black line). The position of the center of one of the hydrophilic beads (appearing at the front of the figure and indicated by the black sphere and an arrow) is fixed at a height d above the underlying surface.
- Collective dynamics of dynamics of myosin motor proteins on actin bundles
The cooperative action of many molecular motors is essential for dynamic processes such as cell motility and mitosis. This action can be studied by using motility assays in which the motion of cytoskeletal filaments over a surface coated with motor proteins is tracked. When moving on actin bundles, myosin motors usually exhibit fast directional motion that reflects their tendency to propagate unidirectionally toward the "plus end" of actin filaments. Recently, a motility assay with actin bundles consisting of short filamentous segments with randomly alternating polarities was presented (by Dr. Anne Bernheim from BGU). These actin tracks exhibit bidirectional motion with macroscopically large time intervals (of the order of several seconds) between direction reversals. Analysis of this bidirectional motion reveals that the characteristic reversal time, τrev , does not depend on the size of the moving bundle or on the number of motors, N. This observation contradicts previous theoretical calculations based on a two-state ratchet model, predicting an exponential increase of τrev with N. We presented a modified version of this model that takes into account the elastic energy due to the stretching of the actin track by the myosin II motors. The new model yields a very good quantitative agreement with the experimental results.
(A)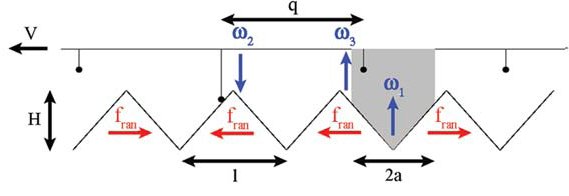
(B)
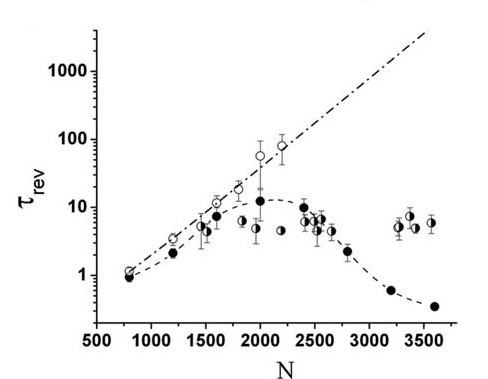
Figure: (A) The model: N point particles (representing the motors) are connected to a rigid rod with equal spacing q. The motors interact with the actin track via a periodic, symmetric, saw-tooth potential with period l and height H. In each periodic unit, there is a random force of size fran, pointing either to the right or to the left (red arrows). The motors are subject to these forces only if connected to the track. The detachment rate ω1 is localized in the shaded area of length 2a < l, while the attachment rate ω2 is located outside of this region. The off rate ω3 is permitted only outside the grey shaded area. (B) The mean reversal time, τrev , as a function of the number of motors N, computed for different realizations of random, a-polar, tracks. The results of our model, which are denoted by solid circles, show very little small dependence of τrev on N, in agreement with the experimental results (half-filled circles). The open circles show the results of a previously proposed model, where the effect of actin stretching is neglected, and τrev grows exponentially with N (dash-dotted line).
My research combines analytical and computational methods of statistical mechanics to understand the physical
principles
behind the functioning of complex biological and biomedical systems. Specific projects in my group focus on the following biophysical system:
Complexes of lipids and DNA molecules.
Supported membranes with reconstituted proteins.
Non-equilibrium dynamics of myosin motor proteins on actin
bundles.